Introduction
Le rapport entre densité, densification urbaine et biodiversité questionne tant les professionnels praticiens de la planification et de l’aménagement urbain que les écologues, les élus ainsi que la société civile. En mai 2019, le rapport de la plateforme intergouvernementale scientifique et politique sur la biodiversité (IPBES), confirmait le besoin de mieux connaître et comprendre les impacts du développement urbain sur la biodiversité, appelant un travail de recherche capable d’orienter l’action des acteurs de la ville 1. Les travaux de recherches menés dans cette direction ont imposé deux concepts clés : le land sparing (LSP) et le land sharing (LSH). Ces deux notions, avant d’être adoptées dans les réflexions sur les modèles de développement urbain et leurs impacts sur la biodiversité, trouvent leurs origines dans la recherche s’intéressant à la cohabitation des modèles d’exploitations agricoles avec la conservation de la biodiversité (Green et al., 2005).
1 A l’image du programme de recherche Baum (Biodiversité, aménagement urbain et morphologie) du PUCA, qui s’est donné en 2019 l’objectif, d’explorer les liens entre densité, formes urbaines et biodiversité. La question mise en avant par le programme était la suivante : « Comment concilier densification du bâti, organisation des constructions et déploiement dans la matrice construite de la ville, d’un réseau d’espaces à caractère naturel, propice à l’accueil de la biodiversité la plus riche possible ».
Transposé aux enjeux relatifs au développement urbain et la biodiversité : le land sparing est associé à une idée de densité urbaine forte, pour contenir le développement urbain dans une compacité qui lui permette d’épargner au maximum les espaces naturels, afin de conserver les milieux et la biodiversité spécifique et fonctionnelle qu’ils accueillent. Le land sharing est pour sa part associé une idée de ville diffuse, permettant l’interpénétration et la cohabitation de l’urbain et des milieux naturels, de leurs espèces et de leurs dynamiques (Lin & Fuller, 2013).
Partant d’un corpus de travaux de recherche empirique, cette méta-analyse a pour objectif d’apporter des éléments de réponses aux questions suivantes : Faut-il densifier les villes par la mobilisation des jardins existants ou bien faut-il mettre les jardins « sous cloche » pour limiter l’effondrement de la biodiversité spécifique et écosystémique constatée à l’échelle mondiale ? Faut-il choisir entre un développement urbain selon un principe de LSP ou de LSH (Youngsteadt et al., 2023) ? Est-il possible de conjuguer LSP et LSH, ou autrement dit, est-il possible de densifier la ville en renforçant sa biodiversité ? (Suarez, 2023)
1 Land sparing et préservation de la biodiversité : si la densité n’est pas l’ennemie de la biodiversité la densification ne l’est pas non plus
La densité urbaine permet un usage compact et rationnel de l’espace. En intensifiant les usages et services que les villes sont capables de rendre aux sociétés humaines, la densité s’oppose au phénomène d’étalement urbain qui implique la dégradation par artificialisation et la destruction de milieux naturels et par extension des espèces qu’ils accueillent (Ren et al., 2023).
En France, les travaux de l’unité d’appui et de recherche PatriNat (OFB-MNHN-CNRS-IRD) sur le croisement des enjeux de biodiversité et des pressions pour l’évaluation du réseau d’aires protégées métropolitain terrestre des sources de pressions menaçant la biodiversité (Léonard, 2020), (Cherrier, 2021), (Suarez, 2023), précisent bien la nature de l’enjeu relatif au développement urbain par rapport à la biodiversité :
Le groupement d’expert n’identifie pas de points chauds de biodiversité dans les environnements urbains et ce quel que soit leur niveau de densité. La notion de points chauds fait référence à la définition du rapport (Léonard, 2020): des zones à conserver prioritairement, car irremplaçables du fait de l’assemblage d’espèces qu’elles abritent, indépendamment de leur niveau de dégradation.
Le risque de pression de l’urbanisation sur la biodiversité relève exclusivement du risque d’extension des surfaces urbanisées sur les milieux naturels (Suarez, 2023).
A l’échelle nationale c’est donc la stratégie du land sparing apparaît comme prioritaire pour réduire l’impact des activités humaines sur l’érosion de la biodiversité et tout particulièrement sur les deux principales sources de pressions sur la biodiversité que sont l’agriculture intensive et l’extension des villes. Rapportée aux enjeux urbains, cette stratégie implique de concentrer le développement urbain en densification, dans les secteurs déjà bâtis et artificialisés, en cohérence avec la direction définie par la loi ZAN du 20 juillet 2023.
Pour mettre en œuvre la stratégie de land sparing en matière de développement urbain nécessaire au respect de la loi ZAN, les secteurs déjà bâtis et artificialisés qui sont aujourd’hui peu denses constituent une ressource clé, notamment la production de logements. La production de ces secteurs peu denses aux cours des dernières décennies a été portée principalement par le développement massif de zones d’habitats individuels, disposant de grands jardins. Ces grands jardins ont ainsi été un des principaux facteurs de l’extension de villes. Mais, alors qu’ils constituent aujourd’hui le gisement principal de foncier pour stopper l’étalement urbain et préserver la biodiversité, on observe dans de nombreux documents d’urbanisme une tendance de préservation, empêchant la mise en oeuvre d’un développement urbain en densification.
2 Le renforcement de la biodiversité en ville grâce au land sharing : 13 travaux de recherche empirique* sur les liens entre densité, densification et biodiversité
2.1 la densification peut contribuer au renforcement de la biodiversité urbaine, notamment en augmentant le linéaire de haies
Varet et al. (Varet et al., 2013) ont conduit une étude visant à mesurer l’impact de la densification sur la biodiversité urbaine en étudiant deux taxons d’arthropodes (araignées et coléoptères carabidés), reconnus pour réagir fortement aux changements des conditions de leur habitat et, à ce titre, souvent utilisés comme bio-indicateurs. En étudiant les assemblages d’arthropodes dans des quartiers pavillonnaires peu denses et dans des quartiers densifiés, les travaux de Varet et al. constatent que les assemblages d’arthopodes sont similaires dans les secteurs densifiés et que l’augmentation du linéaire de haies induit par la densification peut contribuer au maintien d’une certaine biodiversité en ville. Une des conséquences du phénomène de densification des tissus urbains par la subdivision des terrains est l’augmentation du ratio de limites séparatives sur lesquelles des haies sont plantées, principalement dans un objectif de confort et d’intimité vis-à-vis du voisinage. Les haies constituent des ressources pour une grande diversité d’animaux en milieux urbain, elles leur fournissent des abris, des sites de nidification, de la nourriture et constituent des corridors pour leurs mobilités dans l’environnement urbain (Beninde et al., 2015), (Gosling et al., 2016). La parcellisation associée à la densification peut ainsi être à l’origine de la création d’un patchwork de milieux et de lisières entre les différents milieux. Ce type de configuration, identifié comme support pour la biodiversité dans les milieux agricoles, pourrait également supporter davantage de biodiversité en milieux urbains (Statistik, 1997).

2.2 Les gabarits bâtis produits en densification douce sont adaptés à des toits terrasses plus résilients et utiles à la biodiversité en ville
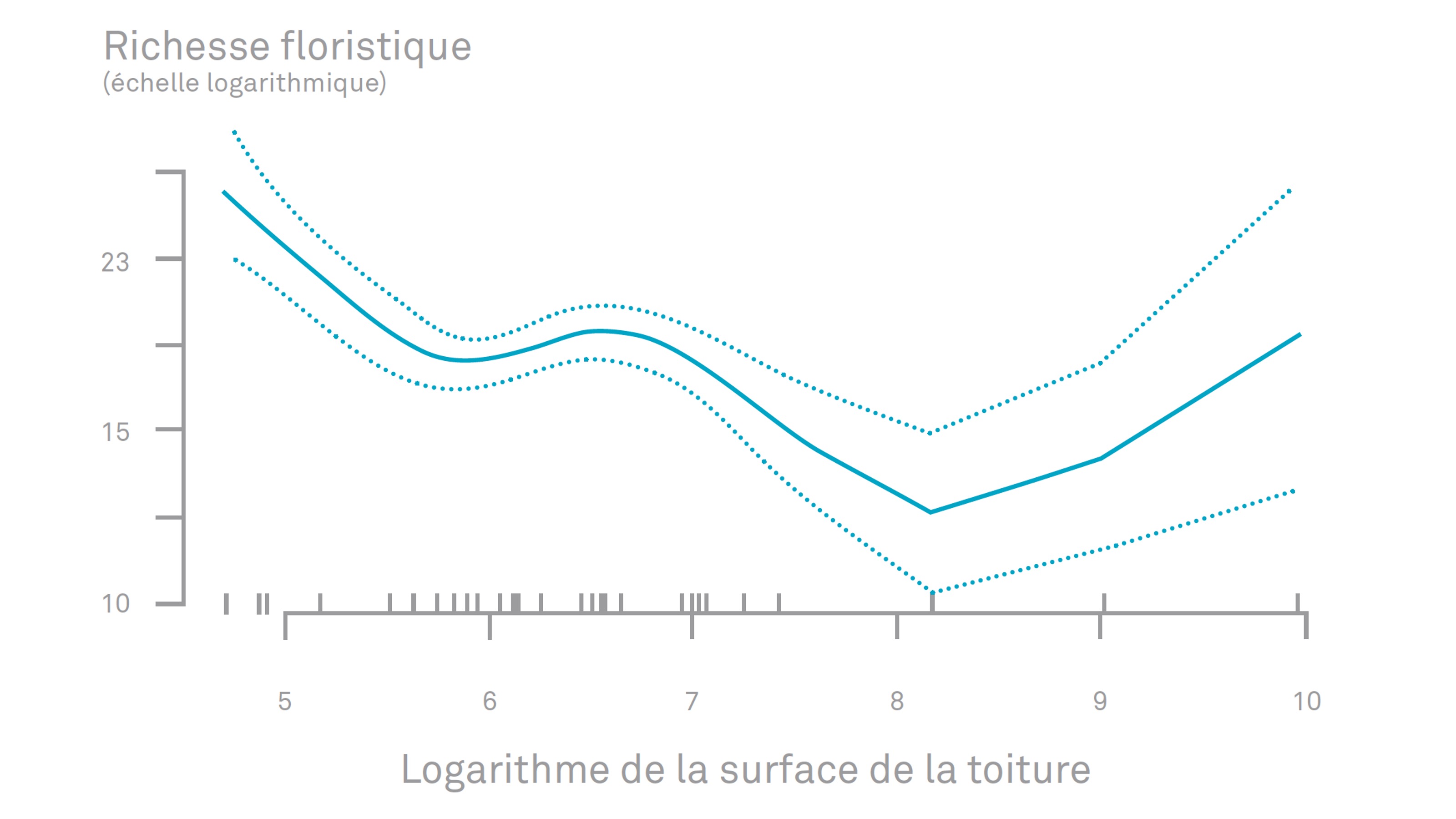
Les travaux du projet de recherche GROOVES sur l’écologie des toitures végétalisées (M. Barra, 2021), menés entre 2018 et 2021 par l’Institut Paris Région et l’Agence Régionale de la Biodiversité sous la conduite de Marc Barra et Hemminki Johan, ont mis en évidence une corrélation négative entre la surface de la toiture et la richesse en plantes spontanées. Pour expliquer ces résultats les chercheurs émettent l’hypothèse que la richesse en biodiversité 2 des toitures terrasses de plus petites surfaces seraient le résultat de leur meilleure résistance aux périodes de fortes chaleurs et à la sècheresse car davantage protégées par les éléments bâtis et naturels avoisinant et donc moins soumis aux vents. La question du gabarit bâti, et en particulier de sa hauteur, est également identifiée comme un facteur à considérer (Maclvor, 2016) : l’étude de la nidification des abeilles sauvages et des guêpes sur 29 toitures végétalisées de Toronto aboutit à la conclusion que les toitures végétalisées installées sur des bâtiments de faible hauteur sont à privilégier pour favoriser ces espèces en milieux urbains (l’intérêt de la toiture pour ces taxons diminuant avec la hauteur, avec un seuil limite identifié à 4 étages d’habitation maximum).
2 La notion de richesse en biodiversité recouvre dans le présent article les notions de richesse spécifique et fonctionnelle, la composante génétique de la biodiversité n’est pas abordée ici.
2.3 La diversité de la palette végétale et la densité urbaine impactent positivement le process de décomposition de la litière organique
Les travaux de Tresh et al. sur l’étude de la capacité des sols des jardins urbains à décomposer la litière organique ont étudié les différents facteurs environnementaux qui impacte ce processus (Tresch et al., 2019). A partir d’un échantillon de 170 jardins urbains situés dans différents contextes de densité dans la ville de Zurich, l’équipe de chercheurs aboutit à deux conclusions :
a) Les pratiques des jardins en matière de gestion et de choix de palette végétale impact très fortement la richesse et la diversité des populations de la faune du sol, éléments clés de sa capacité à décomposer la litière organique.
b) La densité urbaine, en créant un micro climat plus chaud dans le sol, impacte de manière très positive le processus de décomposition de la litière organique en créant un milieu favorable à un développement abondant de la faune du sol (notamment les isopodes). Ces résultats corroborent un corpus d’études antérieures menées en Europe et aux Etats-Unis (Melliger et al., 2017), (Nikula et al., 2010), (Pouyat & Carreiro, 2003).
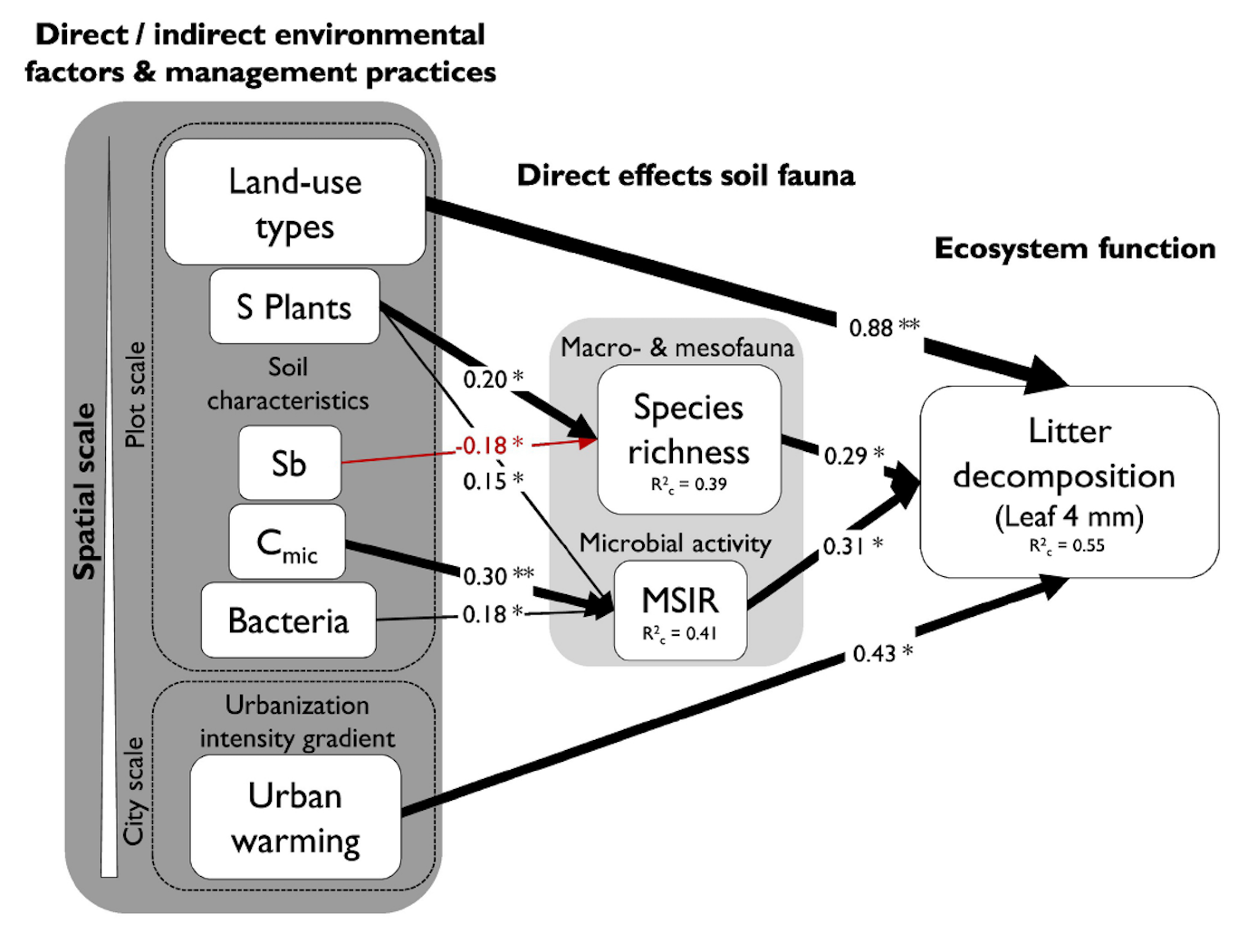

2.4 La richesse des flores présente dans les jardins résidentiels augmente avec la densité urbaine
Knapp et al. (Knapp et al., 2012) ont étudié la phylogénétique, la diversité et la composition des traits fonctionnels de la flore dans les jardins privés, selon le gradient de densité urbaine. En complément, leur étude a cherché à identifier les caractéristiques de la flore spontanée dans le corpus de 137 jardins résidentiels de la métropole de Minneapolis-Saint Paul, en comparant sa phylogénétique et sa composition fonctionnelle par rapport à un espace naturel de référence situé à proximité des sites étudiés (réserve naturelle de Cedar Creek). L’analyse des résultats a établi une corrélation positive entre biodiversité végétale et densité urbaine. L’étude aboutit à la conclusion que les jardins comptent davantage de biodiversité par hectare dans les secteurs densément bâtis que dans les secteurs à faible densité ,alors que la diversité phylogénétique et la composition fonctionnelle des flores ne changent pas avec la densité d’habitation.
2.5 La petite dimension et la forte fragmentation des jardins en milieu urbain dense n’est pas un frein pour qu’ils constituent une ressource clés pour certaines espèces en ville: le cas de la pipistrelle commune à Paris intra-muros
Les travaux de Mimet et al. apportent un éclairage sur la contribution des jardins privés dans un contexte de très forte densité urbaine en s’intéressant aux contributions respectives des grands epaces verts et des jardins privés dans Paris intramuros (Mimet et al., 2020) pour la pipistrelle commune (Pipistrellus pipistrellus).
L’étude établit qu’un réseau de jardins de petite taille peut avoir un rôle plus important que les grands espaces verts publics pour le maintien d’espèces protégées en ville comme la pipistrelle commune. Malgré la fragmentation, les faibles dimensions des surfaces de jardins et la plus faible proportion d’espaces verts privés à Paris (36 % du total des espaces verts), ces derniers contribuent néanmoins à hauteur de 47,9 % à la disponibilité de l’habitat des chauves-souris et diminuent la résistance de la matrice urbaine de 57 %. Les auteurs soulignent que la répartition des jardins privés dans la matrice de la ville et la composition de la végétation de ces zones semblent particulièrement bénéfiques pour la disponibilité et la connectivité de l’habitat des chauves-souris. L’étude met à ce titre en avant le rôle clé des jardiniers et des modes de gestions qu’ils opèrent dans la valeur écologique des jardins pour la faune.
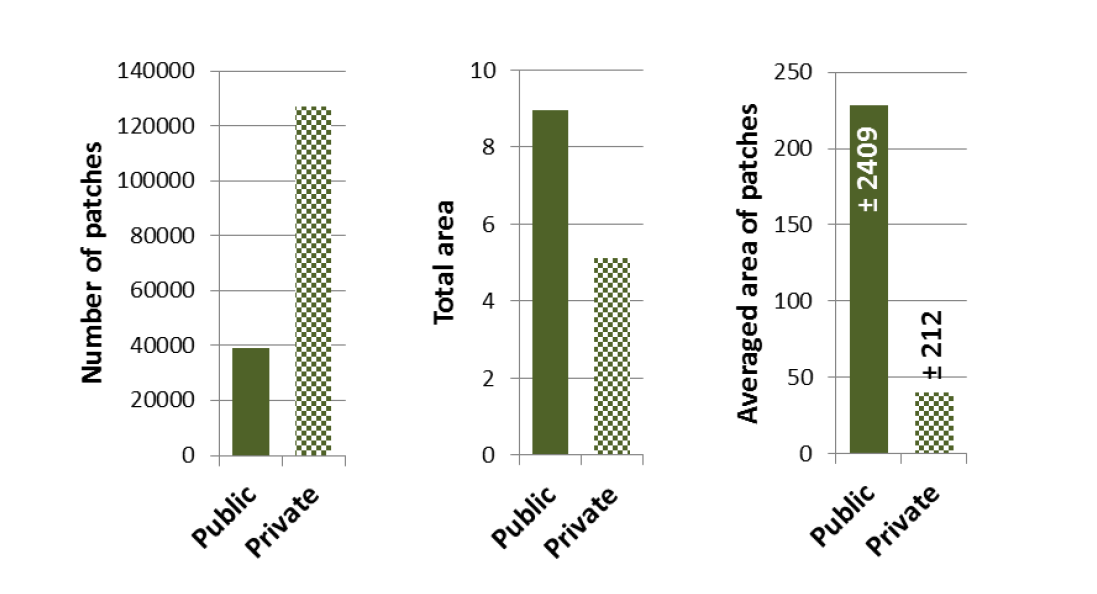
2.6 Les grands jardins se sont pas davantage support à la biodiversité que les petits jardins
Certains travaux de recherches défendent des thèses opposées aux conclusions des travaux mentionnés précédemment (Varet et al., 2013), (Knapp et al., 2012) ou (Mimet et al., 2020). Ils affirment alors que la surface d’un jardin est un facteur déterminant de sa capacité à permettre le renforcement de la biodiversité en ville. Selon cette thèse plus un jardin est grand, plus il fait de place à la nature.
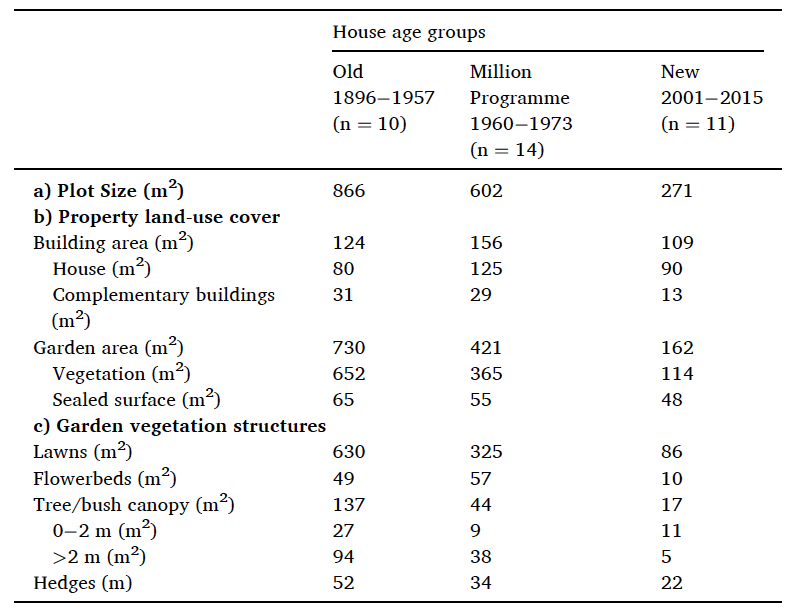
Parmi les récents travaux qui alimentent cette thèse, l’article d’Helena I. Hanson et al. (Hanson et al., 2021) et éclairant : à partir de l’étude de l’usage du sol dans un corpus de 35 jardins privés situés dans la région de Lund en Suède et de la conduite d’entretiens semi-directifs avec leurs propriétaires, les auteurs affirment que la densification urbaine, en réduisant la taille des jardins, réduit également leur biodiversité. Les données issues du travail d’inventaire sur le corpus de jardins fait pourtant ressortir (et les auteurs l’évoque succinctement dans leur conclusion) que plus les jardins sont grands, plus ils sont pauvres en biodiversité car largement occupés par des pelouses (86% de l’occupation du sol dans les parcelles d’une surface supérieure à 730m² contre 53% pour les parcelles de surface inférieure à 271m²). L’analyse de ces données permet pourtant de formuler deux affirmations :
a) Il n’est pas possible de démontrer à partir du corpus que les grands jardins sont plus favorables à la biodiversité : si cette idée est vraie en théorie (les auteurs évoquent au sujet des grands jardins un “potentiel à être support de biodiversité urbaine”), elle se révèle fausse dans la pratique.
b) La taille réduite du jardin est favorable aux usages du sol utiles à la biodiversité. Le fait d’avoir moins de surface à entretenir augmente l’intensité du jardinage et donc la valeur de chaque mètre carré de jardin pour la biodiversité en donnant la capacité aux jardiniers d’adopter des aménagements et des modes de gestion qui lui sont favorables.
Dans une récente méta-analyse (Delahay et al., 2023) défend également la thèse selon laquelle la réduction de la taille des jardins constitue une menace pour la biodiversité en ville. Comme fondement à cette affirmation il met en avant une référence souvent invoquée pour justifier que la richesse en biodiversité d’un jardin est liée à sa grande taille : les travaux du projet de recherche BUGS (Loram et al., 2008), (Davies et al., 2009). L’article de Loram et al., Urban domestic gardens (XII):The richness and composition of the flora in five UK cities (Loram et al., 2008), est souvent mise en avant comme référence sur le sujet de la corrélation entre surface du jardin et biodiversité. Pourtant cette étude ne s’attache pas à démontrer que la biodiversité d’un jardin est relative à sa superficie, elle démontre une corrélation entre la surface de jardin qui n’est pas en pelouse (les surfaces de pelouses sont expressément exclues de l’évaluation) et la biodiversité spécifique végétale.
La déformation de la conclusion de cette étude rends possible un raccourci qui fonde l’idée fausse d’une corrélation directe entre l’étendue d’un jardin et sa richesse en biodiversité. Richard J. Delahay conclut d’ailleurs sa méta-analyse en indiquant que “Si la richesse en espèces des jardins est largement déterminée par ce que les ménages sont capables et choisissent de planter, et si les plantes sont à la base de la contribution des jardins à la biodiversité urbaine, alors toute plantation est importante, quelle que soit la taille ou l’emplacement du jardin.” (Delahay et al., 2023). Une clarification sur ce point dans le corps de l’article aurait pourtant été précieuse, car cette lecture erronée du rapport entre la surface des jardins et la biodiversité urbaine et couramment diffusée parmi les professionnels praticiens de la planification urbaine.
On la retrouve par exemple dans l’étude de l’APUR sur les tissus pavillonnaires d’Ile-de-France, publiée en juin 2023 (Christiane BLANCOT, 2023). Les conclusions de cette étude prônent un statut quo de préservation, contraire à la densification des quartiers pavillonnaires. La justification avancée par l’étude est la suivante : les tissus pavillonnaires ayant les surfaces végétalisées les plus importantes au sein des tissus urbanisés de la Métropole du Grand Paris, ils joueraient un rôle majeur pour l’adaptation aux canicules dans un contexte d’évolution des conditions climatiques. Cette qualité découlant, selon l’étude, du seul taux de végétation, lequel serait mis en péril par toute forme de densification potentielle à venir.
L’étude de l’Apur n’aborde les jardins qu’en tant que surface et principalement sous l’angle de leur rôle potentiel de réduction du phénomène d’îlot de chaleur urbain. La biodiversité urbaine, si elle est évoquée, n’est pas un sujet de développement de l’étude. Pourtant, le rapport pointe lui aussi les limites d’une approche fondée sur l’acceptation générale d’une valeur intrinsèque des jardins en ne considérant que leur superficie, soulignant que « Les jardins des tissus pavillonnaires sont dominés par la présence de pelouses ou gazons. Les gazons participent aux compositions paysagères de nombreux jardins, les « mauvaises herbes » qui y poussent sont souvent arrachées par souci esthétique. Il en résulte un milieu monospécifique, donc sans diversité, et donc un sol écologiquement pauvre inapte à stocker l’eau de ruissellement tout au long de l’année. La résistance des gazons aux épisodes caniculaires est ainsi très faible, les gazons nécessitent une alimentation continue en eau sinon ils dépérissent rapidement quand il fait chaud. Leur présence, en tant que milieu monospécifique, ne permet pas une valorisation optimale du cycle annuel de l’eau, pourtant capitale dans l’adaptation climatique. »


Dans chacun des travaux de Loram et al. ou de l’Apur et de Hanson et al. se retrouve, en filigrane, la question du rôle du jardinier et de l’adéquation entre la surface de jardin qu’il a à entretenir et ses capacités à adopter des aménagements et des modes de gestion favorables à la biodiversité, car la volonté de maintenir l’espace domestique “sous-contrôle” et la contrainte de temps dédié à l’entretien du jardin (Hanson et al., 2021) font souvent de la pelouse le premier choix pour simplifier les actions de maintenance (Frileux, 2010) (Dauphin, 2020).
2.7 L’action du jardinier, levier clé du renforcement de la biodiversité urbaine, est dépendant de l’adéquation des caractéristiques du jardin avec la capacité du jardinier
Les travaux de recherche s’accordent à identifier deux leviers pour l’amélioration de la biodiversité urbaine dans les jardins :
a) L’enrichissement des palettes végétales cultivées dans les jardins et l’adéquation des essences qui les constituent aux conditions de milieux. (Threlfall et al., 2017) (Mata et al., 2023) (Delahay et al., 2023)
b) L’adoption par les jardiniers d’aménagements, de pratiques et de modes de maintenance des jardins favorables à la biodiversité (par exemple la création de point d’eau, de toitures végétalisées, la fréquence des tontes et des tailles, l’absence de recours aux produits phytosanitaires, l’apport régulier de matière organique,…) (Loram et al., 2011) , (Tresch et al., 2019), (Mata et al., 2023), (Tassin de Montaigu & Goulson, 2024),(Delahay et al., 2023).
On notera l’absence de la question de la superfice des jardins comme levier pour agir en faveur de la biodiversité (Delahay et al., 2023).
Le rapport entre a), b) et les caractéristiques des jardins est complexe. Il ne s’agit pas un simple rapport de “causes à effets” et le fondement qui guident les actions des jardiniers dépendent de nombreuses exigences individuelles. Si les caractéristiques physiques du jardin comptent, la question de l’âge du jardinier ou encore la durée de résidence sur le terrain comptent parmi les nombreux critères mis en avant dans différentes études (Loram et al., 2011), (Tresch et al., 2019), (Hanson et al., 2021), (Tassin de Montaigu & Goulson, 2024).
La diversification des palettes végétales et la modification des pratiques de gestion peuvent être misent en pratique sur des jardins de très petites dimensions et avoir des effets puissants en terme de renforcement de la biodiversité en ville. En combinant ces deux leviers sur une micro-parcelle de 195m² dans un environnement urbain dense et soumis à de fortes pressions d’usage à Melbourne, Luis Mata et al. ont démontrés la capacité de ces deux leviers combinés à libérer le potentiel d’accueil de biodiversité des petits espaces jardinés en ville (Mata et al., 2023) . Afin d’objectiver la capacité des actions de végétalisation à atténuer les effets négatifs de l’urbanisation en facilitant le retour d’espèces rares ou disparues du milieu urbain, les chercheurs ont développé et appliqué un protocole de suivi et de collecte de données sur quatre années. L’analyse des données a permis de mesurer une augmentation par un facteur 7 de la richesse en espèces d’insectes par rapport à la situation initiale.
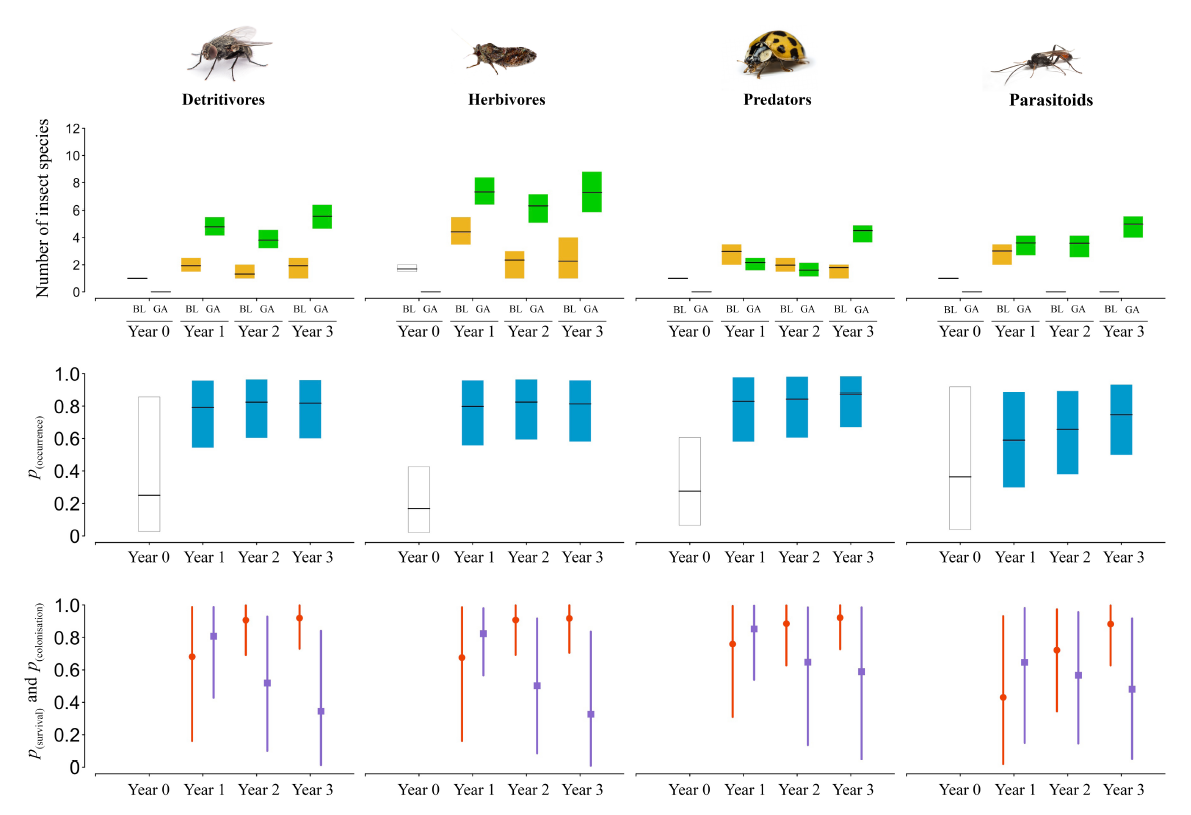
![(a) Réseaux quantitatifs bipartis d’interactions (accords) entre espèces végétales (référence : boîtes blaches ; verdissement [année 3] : vert) et clades d’insectes (détritivores : orange ; herbivores : jaune ; prédateurs : bleu ; parasitoïdes : violet). La largeur de la corde reflète la relative richesse des espèces d’insectes du clade donné représenté dans l’interaction. (b) Au niveau du réseau (richesse des interactions, diversité et uniformité; et spécialisation : H 2) ou au niveau de l’espèce (spécialisation plantes et insectes : d plantes, d insectes) métriques pour l’année 0 (boîte blanche) et Année 1, Année 2 et Année 3 (cases bleues), telles qu’estimées par les modèles linéaires généralisés. Les lignes noires horizontales représentent les estimations moyennes et encadre l’incertitude associée aux intervalles de crédibilité à 95 %. (Mata et al., 2023)](img/mata_2.png)
3 La densification douce : faire du land sparing et du land sharing une seule et même action
3.1 Evolution des jardins pavillonnaires sur 10 ans à Montfermeil : que se passe t-il sur les parcelles qui ne font pas l’objet de projets de densification ?
Dans le cadre du projet de recherche ANR BIMBY (Miet, 2012), un travail d’évaluation des jardins et de l’intérêt de leurs usages du sol pour la biodiversité a été réalisée sur un échantillon de 322 parcelles composant quatre îlots représentatifs des quartiers de Franceville et des Coudreaux en 2013. Il ressortait de cet état des lieux que dans 72% des cas, le jardin se limitaient à des espaces de pelouses et que dans 5% des cas les espaces libres étaient entièrement minéralisés. Seul 23% des parcelles présentait un usage du sol du jardin favorable à la biodiversité (dominante du couvert ligneux et espaces de cultures potagères).
En 2023 un travail d’actualisation des données sur l’usage des sols des jardins sur l’ensemble des parcelles de ces quatre îlots (335 parcelles à l’issue de recompositions cadastrales qui ont eu lieu au cours des dix années) a permis d’évaluer la dynamique d’évolution de l’usage des sols des jardins en dehors de tout projet de densification. Ce travail d’analyse comparative aboutit aux conclusions suivantes :
67% des parcelles étudiées présentent un usage du sol dominé par les pelouses (226 parcelles). Sur la période de 10 ans, cette part reste donc relativement stable.
Les parcelles dont les extérieurs sont dominés ou exclusivement minéralisés ont augmenté de +62% (de 34 parcelles en 2013 à 55 en 2023). La moitié des nouvelles parcelles dépourvues de jardins constatées en 2023 est le fruit d’évolutions spontanées des aménagements par les propriétaires (aucune mutation sur ces propriétés au cours des 10 ans).
Une perte significative du couvert végétal a été observée sur 115 parcelles : 4 fois sur 10 cette perte est liée à une mutation suivie de réaménagements par les nouveaux propriétaires, 6 fois sur 10 à des réaménagements engagés par les propriétaires eux-mêmes. La perte de couvert végétal se traduit pour moitié par l’arrachage de haies ou d’arbustes, pour un quart par la minéralisation de surface de pelouses et pour un quart par l’abattage d’arbres.
Sur la période 2013-2023, la part des jardins dominés par un couvert arboré reste stable (l’augmentation du couvert arboré sur certaines parcelles compensant les pertes observées sur d’autres)
Les espaces à dominante de cultures potagères ont diminué de moitié sur la période et ont été remplacés par des pelouses pour leur majorité et dans une moindre part par des espaces minéralisés.
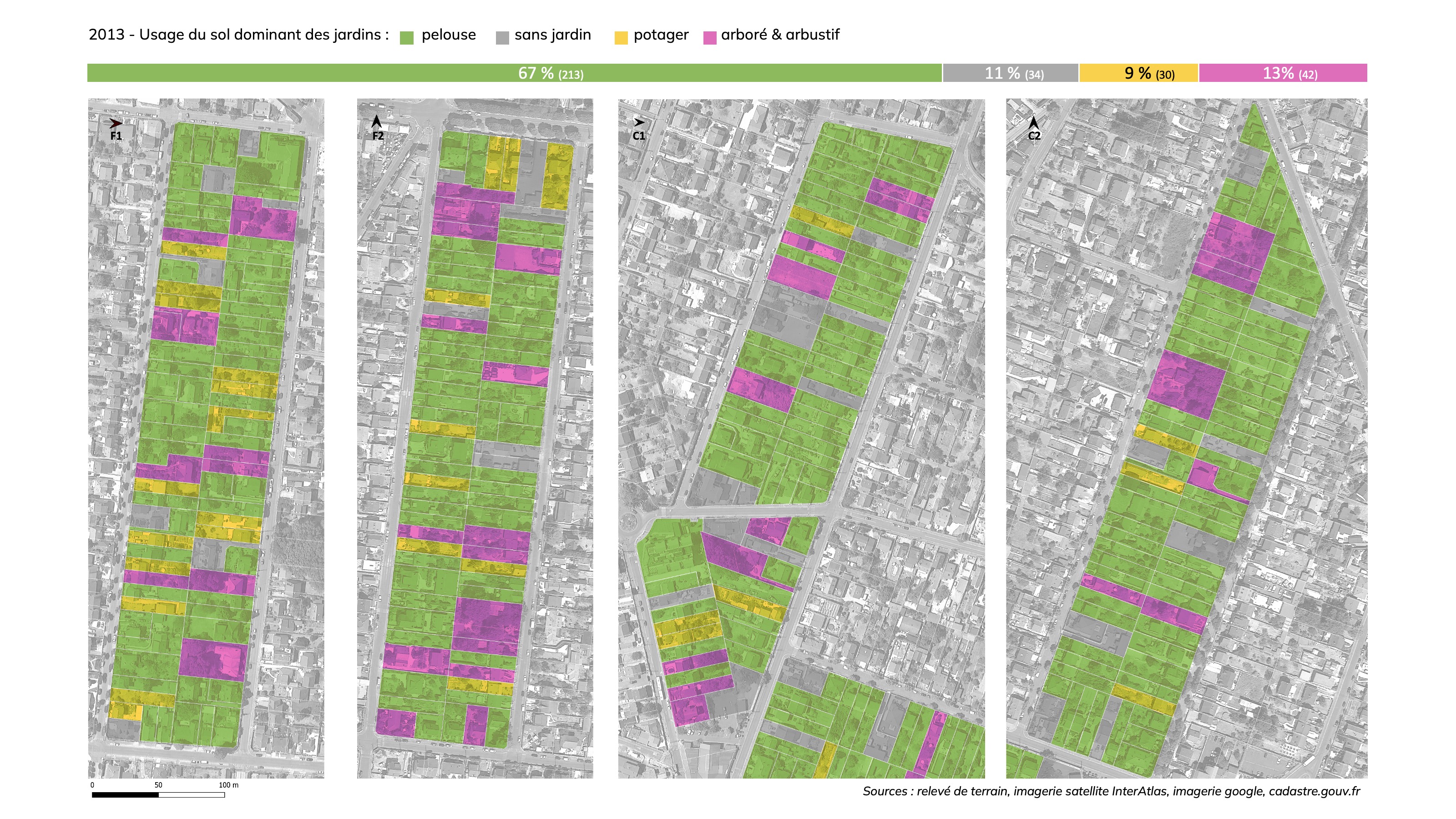
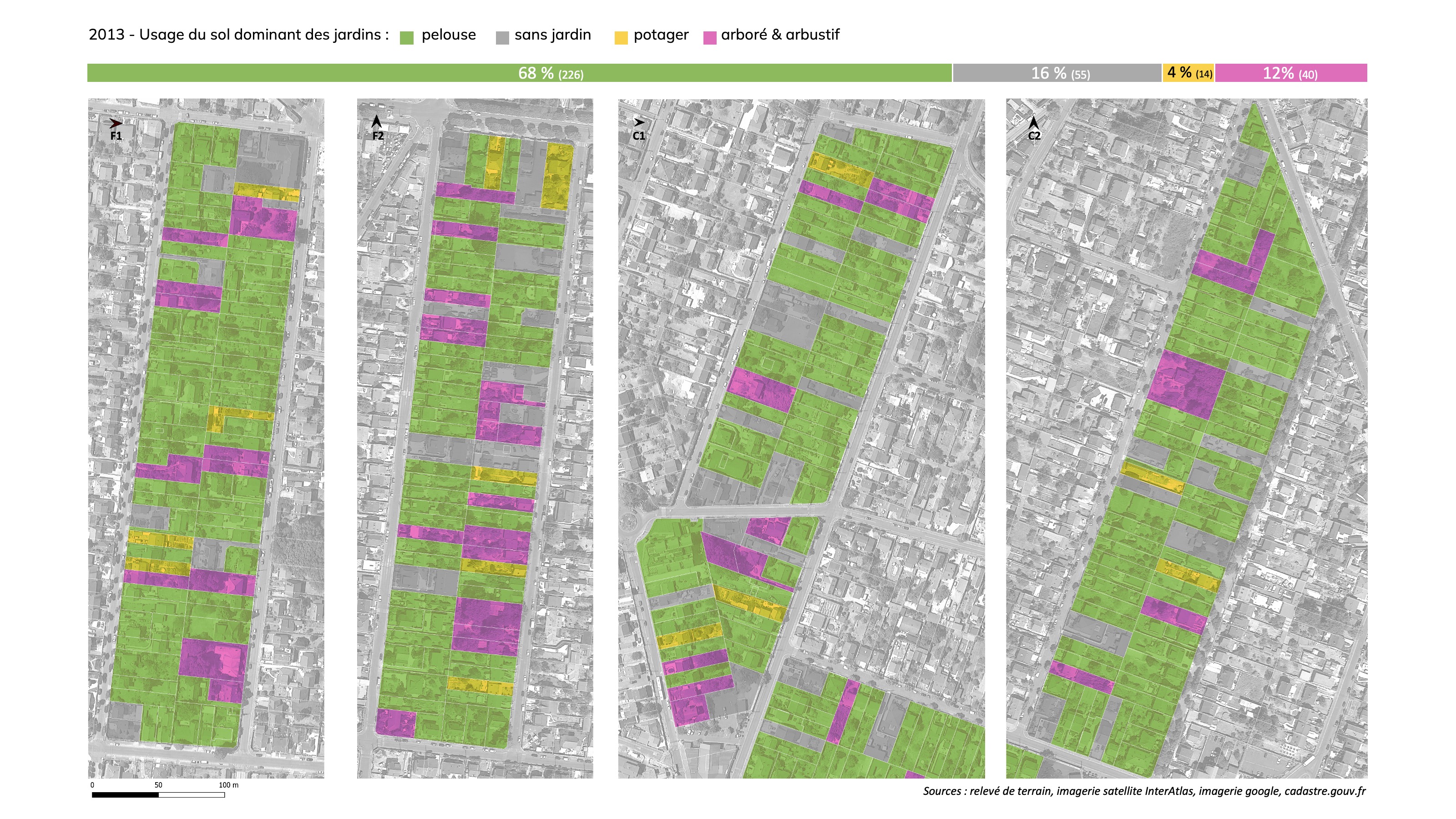
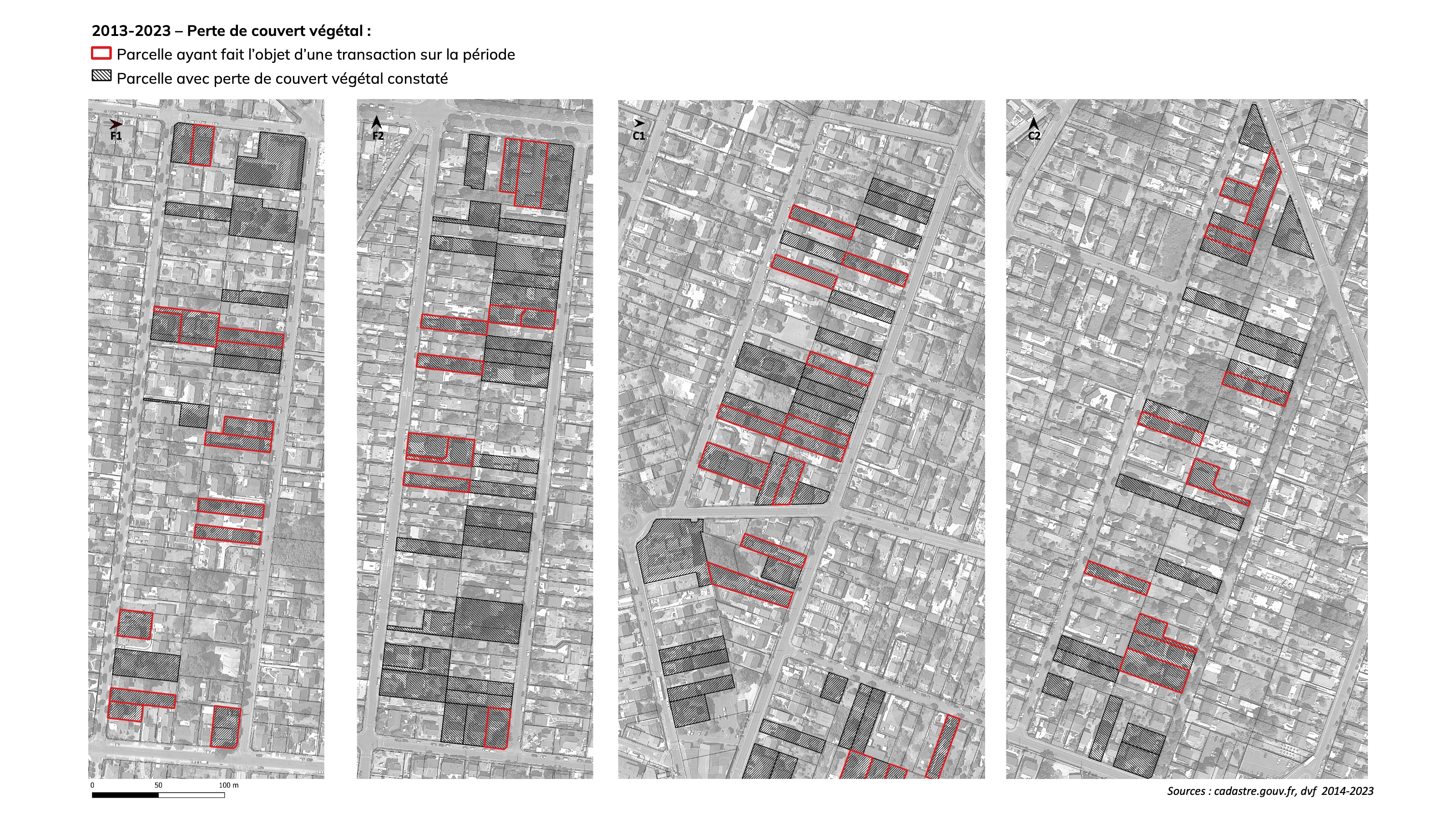
L’analyse comparée de l’usage du sol sur l’échantillon de parcelles à Montfermeil permet d’identifier que la dynamique de perte du couvert végétal et des aménagements favorables à la biodiversité dans les jardins sont majoritairement le fait des propriétaires en place, sans lien avec des projets de densification urbaine. Ce constat appelle d’autres investigations, mais permet de formuler une hypothèse suivante : l’arrachage de haies et d’arbustes, la disparition des potagers et leur remplacement par des surfaces de pelouse, voir des surfaces minéralisées, et plus globalement l’augmentation des surfaces minéralisées semblent traduire un choix des jardiniers pour aller vers une simplification de l’entretien du jardin. Ce choix de la simplification, dans le but de limiter le temps et/ou les dépenses à dédier au jardin, peut résulter d’une dimension de jardin qui n’est pas optimale pour permettre aux jardiniers de s’engager ou de conserver des pratiques de jardinage favorables au maintien et au renforcement du couvert végétal et de la biodiversité.
3.2 La première vertu des projets de densification douce est d’adapter la dimension du jardin à la capacité du jardinier
La densification douce, en redimensionnant les jardins à une taille permettant une intensification du jardinage, rendent possible l’activation des deux leviers d’amélioration de la biodiversité urbaine que sont la diversification et l’enrichissement de la palette végétale et l’adoption de mode de gestion favorable à la biodiversité. Le jardin de Joseph Chauffray à Sotteville-lès-Rouen représente un cas d’école sur l’impact des pratiques et des usages du jardinier en matière d’accueil de biodiversité et de production vivrière à l’échelle domestique en milieu urbain.
La question de l’adéquation de la taille du jardin avec le déploiement d’aménagements, de plantations et de modes de gestion adaptés est au coeur de cet exemple éclairant sur la capacité du jardinier à avoir un impact en matière de biodiversité urbaine.
Le jardin de Joseph Chauffray est situé dans un tissu ancien de lot libre dense sur une micro-parcelle de 255 m² environ. Le jardin occupe une surface de 150m², incluant la terrasse et les allées. Lorsque l’actuel propriétaire acquiert la parcelle au début des années 2010, le jardin présente les aménagements classiques d’un jardin pavillonnaire. En dehors de la terrasse et des allées, on ne trouve aucun arbre. Les arbustes se limitent à des thuyas et des lauriers du Portugal taillés en topiaires géométriques, quelques hortensias et rosiers et un troène. La richesse de biodiversité végétale est estimée à une quinzaine d’espèces, incluant le couvert de pelouse.
Inspiré par le concept de permaculture de David Holmgren (Holmgren, 2002), Joseph Chauffray décide de faire table rase : son objectif avec ce jardin et de découvrir jusqu’où il est capable d’aller en termes autonomie alimentaire en empruntant l’approche de la permaculture (pas de travail du sol, pratique réparatrice des écosystèmes). Poursuivant de façon continue expérimentation et optimisation d’un système de gestion du jardin, Joseph Chauffray aboutit à un exemple frappant du potentiel des petits jardins urbains en termes de biodiversité, mais aussi de simplification des actions de maintenance et de productivité nourricière. La seule pratique du jardinier a complètement transformé le jardin et la documentation précise dressée par Joseph Chauffray au fil des années permet de mesurer son impact : la biodiversité spécifique y a été multiplié par 15 (de 15 à 226 espèces végétales) et le jardin produit plus de 300kg de fruits et légumes par an. Il consacre en moyenne 3 heures par semaine à ses 30 m² de potager et à ses 15m² de surface dédiées aux cultures fruitières, le reste du jardin étant occupé par la terrasse, les allées, une annexe faisant fonction d’atelier et de lieu de stockage et d’un bassin d’agrément. (http://josephchauffrey.fr/visiter-le-jardin/)


Dans une interview qu’il donnait en 2015 pour le média web BASTA, Joseph Chauffrey donnait les clés de cette incroyable transformation, rappelant le caractère essentiel de la petite taille du jardin dans cette réussite : « Une des clés pour augmenter la productivité du jardin (en permaculture), c’est de réaliser un travail très soigné, sur tout le jardin. Pour cela, il faut passer beaucoup de temps et ce n’est possible que parce que mon jardin est petit.»
La question de l’intensité de jardinage et donc de la dimension du jardin avec l’approche de la permaculture est également soulevée par les travaux de Goy et Bollinger qui indiquent que, si à surface équivalente, il serait possible de produire 5 à 10 fois plus d’énergie alimentaire au moyen de la permaculture qu’au moyen des techniques actuelles de cultures potagères, celle-ci demande davantage de main d’oeuvre et n’est principalement mise en pratique que sur des surfaces réduites (Goy & Bollinger, 2020).
Réutilisation
Citation
@online{hanss2024,
author = {Hanss, Thomas and Miet, David},
title = {La densification peut-elle être un levier pour améliorer la
biodiversité\,?},
date = {2024-01-11},
url = {https://publications.vv.codes/densification-biodiversite.html},
langid = {fr}
}